Introduction
Our study of Eastern North American prehistory began in 2010 with caribou range reconstruction of southwestern Ontario and the adjacent states of New York and Michigan. This successful model showing overlapping herd ranges and hunting territories was based on a dozen field seasons surveying, locating and excavating 1004 Canadian Barrenland archaeological sites where hunters from the 8000 year-old1 Plano period to the Present relied on caribou. Using artifacts from C14-dated levels involving four very different traditions, we were able to assign surface sites to phase. We needed to do this to investigate long-term use of each water-crossing.
 Central Part of the Appalachian Chain |
 Thelon Herd in distance |
Seasonal herd observations by field crews and wildlife biologists during aerial surveys, plus past sightings by Hearne in the 1770s and later explorers and canoeists permitted the start of herd range reconstruction. We found that the 1970s and 80s Beverly herd range borders were conservatively maintained for several centuries.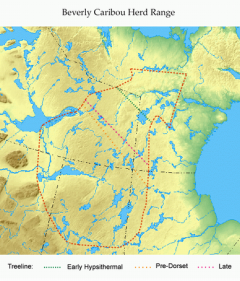 We then tested hunter territorial continuity over time by flying, canoeing and walking well outside the known Beverly range. We surveyed up to the Back River to the north, Artillery Lake to the west, the Dubawnt River to the east and Churchill River to the south. Over a dozen years we found 1004 sites with no sites outside the range. This confirmed an overlap of hunting territory and herd range for each single band for 8000 years. Their borders remained stable throughout very warm postglacial times and the very cold 1500-700 BC Pre-Dorset Inuit-like period.
We then tested hunter territorial continuity over time by flying, canoeing and walking well outside the known Beverly range. We surveyed up to the Back River to the north, Artillery Lake to the west, the Dubawnt River to the east and Churchill River to the south. Over a dozen years we found 1004 sites with no sites outside the range. This confirmed an overlap of hunting territory and herd range for each single band for 8000 years. Their borders remained stable throughout very warm postglacial times and the very cold 1500-700 BC Pre-Dorset Inuit-like period.
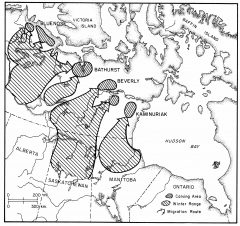
Barrenland Herd Ranges
We found that hunting territory and herd range overlapped in Ice Age France with reindeer, an interbreedable cousin of caribou. Using camp seasonality determined by measuring reindeer tooth cementum increments, I tracked Magdalenians hunting reindeer 17,000 years ago (17K) throughout the country, reconstructing a half-dozen herd ranges and hunting territories. Unsatisfied with two regions and wanting more proof, I did the same in Mesolithic northern Soviet Union (later Russia) using metric and non-metric comparison of blades and other tools, knowing there should be shorter worn conserved ones in the herd’s winter range because toolstone sources were distant and snow-covered, like I found in the Barrens.
While working in the Northern Hemisphere for several dozen years, I found that Eastern North American (ENA) archaeologists had reached an impasse interpreting widespread usage of their Palaeoindian (PI) sites. Some had caribou remains but the many that didn’t were thought by their investigators to represent caribou hunting. After proving overlap of ranges and territories in the three above regions, I thought I could help ENA archaeologists using a broader approach to their huge area. This required knowledge of herd and hunter behavior throughout the year, something that GIS tools could help bring a new perspective on.
We began by leveraging GIS tools in Ontario and bordering States. We had access to Ontario PI fluted point locations which we compared with modeled glacial lake margins and surface vegetation during the advance and retreat of the Ice Sheet. In order to develop and fine tune GIS models of caribou migration corridors for Ontario we tested our simulations against known sites and mapped migration paths in the Barrenlands. We found Barrenland site locations overlapped with the GIS simulated migration caribou routes, confirming that our Ontario model might be expanded to the Upper Great Lakes and northern New England from whence Ontario PI had come. It became a matter of comparing forage zones and archaeological site locations.
Another method of investigation involved connecting sites with toolstone sources. Lines of sites and toolstone routes from Ontario continued into the Upper Great Lake States and New England and then into the Maritimes, their locations heavily influenced by glacial lake, river and ocean shores, and mountains. More research showed the pattern continued deep into the Appalachians, north to Labrador and south to Alabama; basically the rest of ENA. But we needed to see if all sites, especially those with fluted point and Early Archaic points that were further north, were hunting sites based on a meat diet, with emphasis on caribou because of its great adaptation to cold and difficult terrain. To do this we had to evaluate the role of caribou in the diet. This was a no-brainer in the Barrenlands where viable alternate game was missing. Not so for ENA, where most archaeologists describe its early inhabitants as hunter-foragers, but based on little evidence, just the finding of a few seeds and fish bones.
Papers on PI diet were absent but had an underlying disposition to caribou in northern ENA, where most caribou remains were found and where living herds still existed. Absence of papers was due somewhat to heavy criticism: “No body; no Murder” or “No caribou; Forget It”. We countered this with two theories: (1) we know that hunters moved north from the movement north of fluted points and they needed game to follow; i.e. dual mobility. This ruled out most fish and plants, leaving birds and megafauna. No culture relies greatly on birds. Mastodons were spruce forest dwellers while mammoths were steppe dwellers. All were unsustainable throughout the year. So the advantage of caribou is that you can follow large herds and consistently have a solid food source available, whereas seasonality greatly limits the availability of other resources and many resources were available only in smaller amounts. Megafauna were also not viable for long periods from the Southeast to the Great Lakes and Labrador, and hunters needed a game that adjusted to boreal forest, taiga and tundra. This game had to be dense and widespread; easily adapting to a wide range of temperature and forage. In addition, it had to be communally hunted; and (2) a sign that it had to be continuously in ENA well before and during PI times, only to quickly disappear thereafter.
We began not with archaeological data, but with palaeontological finds because there is little evidence of caribou in the archaeological record, but there are areas where caribou remains are present in the paleontological record, in Appalachian caves and offshore near the Continental Shelf. Cave exploration finds by Guilday, Parmalee and others near Appalachian summits were crucial because caribou prefer high isolated calving grounds where pregnant cows safely calve on open tundra where bears and wolves are usually avoidable or where their ranges are separate. Nature plays a role here because the herds’ chief predator, the wolf, dens with pups in the valley when cows calve at elevation. How then do we explain caribou finds in summit caves?
Hunters were absent and usually did not live in caves anyway. Bears are the most common calving ground predator for Ungava’s George River herd which descended from Appalachian herds. In fact, bears may be the main predator because wolves den with pups when herds are calving. Cave bears were present from Alaska to Mississippi, and they became extinct about 11.6K during PI times. But it was more likely that other bears and smaller carnivores that exist today in the Appalachians dragged caribou carcasses into their dens to be consumed.
In northwestern Georgia, caribou remains are in Yarbrough Cave with those of black bear, weasel, otter, cougar and bobcat. Caribou remains are also in Bell Cave in northern Alabama; Charleston in South Carolina; and in Tennessee in the Baker Bluff (with black bear, jaguar and fox), Beartown and Guy Wilson Caves. These caribou remains do not include antler, and post-partum cows quickly lose their antlers. Bears prey on the George River cows while the mottled brown and tan fur of their calves blend with immature tundra vegetation, upon which they lie motionless. Calves are also odourless, while the easily seen, moving cows are pursued right past their inert calves (pers. comm., Tom Bergerud, March, 2017). Post-partum cows were possibly blood-scented and bears have a much better sense of smell than eyesight. I suggest the same thing happened in Appalachian calving grounds, where cows would be reluctant to stray far from their calves. Ultimately, a bear or other carnivore killed and dragged their carcasses into a cave to be consumed without having to defend its meal from other carnivores.
The remoteness and relative freedom from predators make calving grounds the most stable part of a herd range. . In the Appalachians they date from 23K to post-glacial forestation when they became unsafe due to inroads of forest wolves and bears. About that time, tundra, taiga and boreal forest forage for herds was also moving north. This biome change and calving ground abandonment caused a permanent range move north. We had to show that hunters followed herds and this depended upon showing that caribou was their major game. Herd dependency was tricky to determine because most archaeologists either didn’t have bone remains in their sites; calling their occupants foragers. Or they assumed they hunted mastodon and mammoth (Proboscidea) like Clovis hunters out West and Pre-Clovis in the Midwest. After reading many papers, we found that the number of confirmed Proboscidean kills in ENA could be counted on one hand.
I was in a quandary because PI needed reliable and permanent game and nothing viable was proven in the literature. I knew caribou:
- was available long before human occupation;
- was the most important big game in cold environments throughout the world;
- was easily harvested in great numbers;
- by-products like the warmest fur and finest sewing sinew were more useful than other game;
- was predictable in its seasonal movements, especially at water-crossings;
- was prolific in reproduction, permitting communal hunting and surplus storage of dried meat and fat needed for over-wintering.
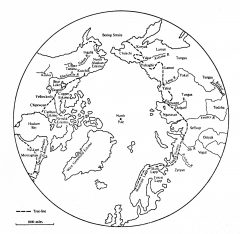 Circumpolar Rangifer Hunters |
 Women with Babies |
Caribou bone was even used for shelters.
 Nadlok shelters – aerial view |
 Nadlok antler shelter – artist rendering |
To ignore these qualities by saying nothing about possible game was avoiding the issue. I had reached the point where we needed to know if the land as far south as Alabama had suitable forage for caribou. I knew its remains occurred even in Mississippi. I knew its calving grounds were down the Appalachian spine when humans were there on its lower flanks. From forage reconstruction I inferred herds descended in their seasonal migrations towards the Mississippi River to the west and the Atlantic Ocean to the east. I knew its remains occur in a continuous line of States on both sides of the Appalachians from Georgia and the banks of the lower Mississippi River north to the Upper and Lower Great Lakes, New England, the Maritimes and Quebec and northern Labrador (Ungava).
Archaeologically, we knew herds and Clovis and Cumberland hunters moved north with post-glacial warming and hunters likely didn’t adopt herd hunting on their way. Therefore, we believe they had this adaptation from the start. But can we prove this? Despite the missing bone and knowing the above facts, plus some early sites with caribou remains, we wanted to see if the site locations of early hunters followed a pattern north, either in lines or clusters, and compare them with herd ranges based on pollen-based caribou forage and GIS-based landforms. This entailed looking for clusters of later fluted points derived from Clovis and Cumberland. From this point on, we modelled hunter sustenance on caribou. We know caribou was on the landscape and was probably a very dominant resource. Through the use of GIS and other models we started to test the theory that sites might be oriented along migration routes. In addition we also explored movement of tool stone to see if that corresponded to herd movement.
We began by looking for a shift in Clovis and Cumberland fluted point locations.
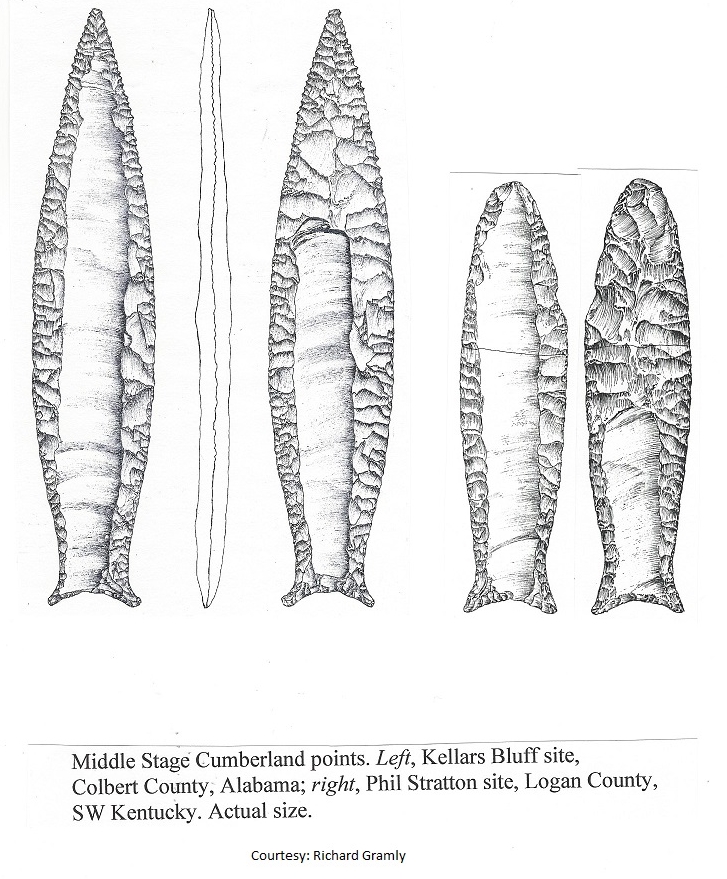

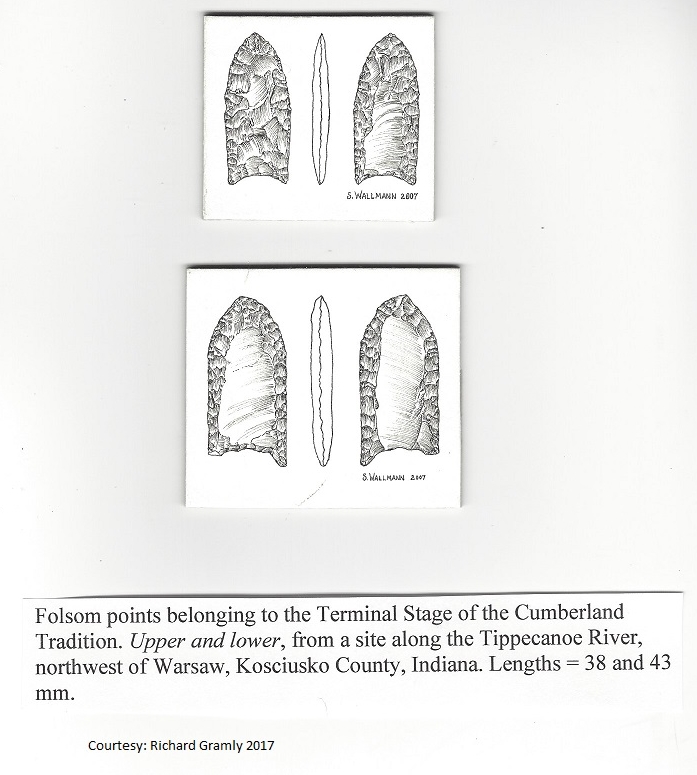
There turned out to be hundreds of them scattered across the Southeast at lower elevations on both sides of the Appalachians, with similar clusters of their derivatives further north. As most herd-harvesting was done at water-crossings throughout the Northern Hemisphere, it was a matter of examining ENA drainage basins and reconstructing them because many were dotted with dams.
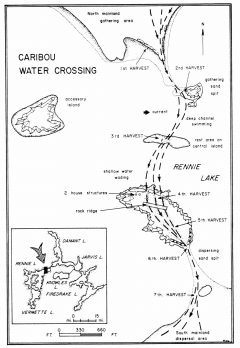
Caribou Water Crossing
Many water-crossings were likely flooded, except those upriver where herds crossed after descending the Appalachians. We needed to reconstruct the ancient shorelines of flooded rivers at their tributaries and at ridges sloping down to their shores.
Most crossings occur where rivers flow at right angles to migration paths, or obliquely where herds crossed where their paths no longer followed the rivers. Wildlife biologists know that a herd’s sense of direction to its calving ground and winter range is keen and it knows where to deviate from an easy river valley to cross a high pass or walk a stony ridge or along a windy insect-free esker to attain its destination. Rivers with narrows, islands and shallows were also excellent fords. Picking a crossing is largely intuitive where herds have long since disappeared.
If a cluster of fluted point locations overlapped a herd range, and if their various daughter styles to the north did the same, I would know I was on the right track. Thus began my search for point locations over a vast area. At first I tried PIDBA maps, but found point locations were county-centric. They were too general for our search for water crossings or a line of sites across the landscape that might indicate a migration route. I then read papers from authors, libraries at my museum and university, but mainly downloadable via e-journals though the university. These totalled about 2500 and were alphabetically arranged by author and duplicated on several hard drives for my core group of three research associates. They were instrumental for checking data while editing our project’s book. Its scope and years of writing left data that needed rechecking. Our papers comprise not only fluted point locations but also palaeoclimatic and pollen data on ancient surface cover, herd and ethnographic hunter behavior, ancient lake and ocean strandlines, Greenlandic ice-core paleo-temperature data, and many maps.
GIS forage and terrain data suggested Southeastern seasonal herd migrations were east-west, unlike those further north where directions were influenced by the Great Lakes, the Maritime mountains and seashore, and very unlike the north-south migrations in the Sub-arctic from Alaska across the Barrenlands to Quebec-Labrador. A time-consuming logistic problem arose when we found site records were confined to state, quite unlike Canada with its Borden national system that even includes Alaska. This meant writing, phoning and visiting state record offices, in addition to checking site reports in papers – a bit of a monumental task but needed. Some states have digitized records. Those that didn’t provided paper copies. They were very kind in this regard.
Before range and hunting territorial reconstruction, we assembled and compared the very diverse and widespread data on palaeoclimate. Pollen charts and interpretations differ from paper to paper according to their analysts’ training. One must also allow for distortion by distant wind-carried pollen, like spruce which can be carried hundreds of km. Spruce is a good sign of herd forage in the form of arboreal lichen. In addition, pollen core locations were very unevenly distributed over ENA and many do not date to fluted point times. As palynologists and palaeontologists also define their finds in Linnaean terms, we used common terms where possible.
Far to the north we examined Ice Sheet margins at different times because they determined the furthest extent of possible herd and hunter habitation. But even thousands of km to the south and even earlier, ice had profound effects on vegetation. Except for icy or snow-covered Appalachian summits, I doubt fluted points hunters actually witnessed ice. The Ice Sheet edge was covered in debris from which flowed meltwater rivers that formed large glacial lakes without fish which PI had no reason to use. Some readers may be surprised that ice advanced almost halfway across North America during its late glacial maximum (LGM), almost to Kentucky, and that caribou were in Mississippi. More subtle are its temperature oscillations from the far north from ice readvance, with alternating tundra and spruce forest to the north. A belt of taiga from the Atlantic to the Mississippi remained across the Southeast for thousands of years, providing refuge for herds and hunters. Range and territory were stalled during the Younger Dryas (YD) cold period. This delay may be simultaneous with the appearance of many fluted point styles in distinct hunting ranges.
About 18K, a belt of boreal forest extended from southern Georgia across northern Alabama and Mississippi to Arkansas. Alongside and to the north, a belt of taiga traced the same east-west path, reaching north into the Carolinas on the east slope of the Appalachians and across western Kentucky to the Indiana border on its western slope. A thin belt of taiga connected these extensions by following the west slope from the Virginias southwest along the east Tennessee border to join the east-west belt in northwest Georgia (see Don Gribble’s separate Paleoclimate presentation). These belts fully enclosed an oval tundra ring along the southern Appalachian spine, with a central core of summit snowfields. Beyond the outer taiga belt, tundra extended from the Ice Sheet southwest to Alabama and northeast to the Maritime Provinces. It continued over southern New England and south of the Great Lakes in an immense U-shaped curve to the Mississippi headwaters in Minnesota. Its northern limit at the Ice Sheet was also the limit for living things, with the likelihood of the presence of caribou and possibly hunters south of its meltwater lakes.
A millennium later at 17K, the biomes have not substantially changed. But at 16K, the boreal forest belt had advanced from South Carolina across northern Georgia, Alabama and Mississippi to Arkansas. Ice had left the coasts of Maine and the Maritime Provinces across New England and south of the Upper Great Lakes. Snowfields persisted along the Appalachian spine enclosed on all sides by tundra. By 15K, the southern taiga belt remained quite stable, suggesting caribou forage was still available. Appalachian snowfields persisted within a circle of tundra, separated only by the thin earlier boreal forest belt. The first Glacial Great Lake Maumee, later to become Lake Erie, appeared as glacial meltwater. To the east, ice lobes extended south into what became the Finger Lakes and Glacial Lake Champlain. By 15K, most of Glacial Lake Maumee appeared, but the east-west taiga belt to the south that had persisted since 18K remained quite stationary. This likely prolonged the presence of caribou and fluted point hunters across North Carolina and the northern borders of Georgia, Alabama and Mississippi. It might also explain the retention of certain fluted point styles.
At 14K, the boreal forest belt was of almost identical width from the Atlantic, across the Carolinas, and northern Georgia, Alabama and Mississippi to Arkansas. To the north, the southern tip of Glacial Lake Michigan appeared. Tundra persisted along the Appalachian spine, but snowfields were seasonal and not permanent. This relict tundra was used by caribou for calving. The Finger Lakes extensions were at a maximum, while the Maine Coast was ice-free. At 13.5K, more of Glacial Lakes Michigan and Maumee were visible, while the boreal forest belt far to the south crept a little further north, but was still in Tennessee, northern Georgia and the Carolinas. At 12.5K, the continuous boreal forest belt widened, its east end reaching Virginia and Maryland. Permanent snowfields and ice were in the mountains of Maine, but the Gulf of St. Lawrence was ice-free, while Newfoundland remained ice-bound.
By 12K, the continuous belt of boreal forest was very irregular, jutting north midway almost to Glacial Lake Maumee and at its eastern end into southeast Pennsylvania, Rhode Island and Connecticut. Taiga with tundra north alongside the Ice Sheet was present from Wisconsin east around Lake Michigan to Illinois, Indiana and southern Michigan to Ohio, Pennsylvania and New York. The mountains of Maine around Munsungan and Newfoundland remained ice-bound. Permanent Appalachian snowfields had disappeared. By 11K, all of the Glacial Great Lakes except for their northern shores were bordered by tundra and taiga. The boreal forest belt had widened substantially, leaving the northern border of Tennessee and North Carolina, and occupying eastern Kentucky and Virginia. Tundra persisted in the Appalachians of Pennsylvania and along the West Virginia-Virginia border.
Lake Agassiz in Manitoba was large and draining into the Mississippi River. By 9.5K, the Ice Sheet was well north of the Great Lakes, with Lake Agassiz to the west now very large. Newfoundland and Labrador were ice-free, as was the North Shore of the St. Lawrence except for a small bottleneck around Sept-Îles. Prince Edward Island had now separated from the mainland. At 9K, the Great Lakes were separated from Glacial Lakes Barlow and Ojibway at the ice edge by thin continuous east-west belts of tundra and boreal forest. These lakes were continuous at times with immense Lake Agassiz next to the Ice Sheet, the latter retreating to central Quebec.
Herds and hunters could follow the North Shore into interior northern Labrador where both date to 7K. Tundra was still present in the mountains of Maine where caribou calved. At 8K, boreal forest surrounded the Great Lakes, except for a taiga border along the north shore of Lake Superior. Tundra had all but disappeared except inland from the North Shore and near the much reduced Ice Sheet, its meltwater forming a gigantic arc of Agassiz, Ojibway and Barlow glacial lakes. By 7K, a thin belt of tundra surrounded the south and east sides of the disintegrating ice, its east and north sides alongside Glacial Lake Tyrrell (Hudson Bay) and Hudson Strait. Lake Agassiz had drained and a cluster of boreal forest remained in Maine and New Brunswick. By 6K, conditions were much like today, with ice disappearance at 5.5K.
The changing biomes previously discussed determined where suitable forage existed for herds. Caribou is one of the most adaptable animals in the world, and this adaptability not only allowed it to thrive wherever suitable forage existed in tundra, taiga and even in deciduous forest to a certain extent. It could even survive on rocky tundra calving grounds where ground and rock lichen and early spring plant shoots were available, and later when warming allowed soil development with browse like willow and many shrubs. The best nutrition was arboreal lichen on mainly spruce but also on other conifers. Some adaptation succeeded in broad-leafed forest where animals browsed on shrubs. This wide range of acceptable biomes allowed herds to move north with forage movement, but also to remain in those biomes where browse remained for centuries in south and central Appalachia and in NEM during the colder YD when tundra and taiga regained former territory.
As aforementioned, biomes comprised largely east-west bands of tundra, taiga and boreal forest stretching from the Atlantic to the Mississippi River, interrupted mainly by the Appalachians, with altitude determining their biomes of snow fields surrounded first by tundra and then by taiga and boreal forest at their base. These upper biomes heavily influenced herd (and hunter) movement north because they were retained longer due to colder temperatures, but also because calving grounds were there, the most conservatively maintained part of the range. With these facts we were able to model herd ranges and their transition north.
Beginning again at 18K, the band of boreal forest stretching from southernmost South Carolina across mid-Alabama and to northern Mississippi and Arkansas supplied caribou with arboreal lichen. It had done so since >36.8K near the North Carolina-Virginia border, well before the LGM. Caribou date 27K in New Bern, North Carolina and 28.3-17.1K at New Trout Cave, West Virginia. The spread of dates in West Virginia suggests lengthy use of the surroundings by herds extending into PI times. Long usage is also in Yarbrough Cave in northwestern Georgia, its 3rd level caribou bone and two molars dating 19-15K, with its 14th level at 22K. In 1991, an amateur spelunker and collector claimed to have found a unifacially-fluted triangular quartz point in the sidewall of the Peccary Room test. Its place in the 18-14K level, plus its matching adhering soil, suggest caribou was hunted by Pre-Clovis and Clovis people in the Southeast. Undated remains are in Kentucky at Big Bone Lick, and dated remains in Tennessee in Guy Wilson, Baker Bluff and Beartown Caves, dating 19.7K, 19.1K, <10.56K and 20K. Linville Cave is an extensive karst system also in Tennessee, where a Clovis point was found not far away.
We mentioned the boreal forest biome, in the vicinity of which many of these finds were made because it was used for a long time, only moving slowly north almost to the Great Lakes when its speed north accelerated. We conclude from this that herds certainly stayed for centuries in the Southeast and its hunters were first pre-Clovis, who likely hunted caribou; then Clovis and Cumberland hunters who formed the roots of these two fluted point traditions. As these biomes moved slowly north, the fluted points did not change appreciably as their bearers crossed the tundra of Tennessee into western Kentucky and over the Carolinas, with the Appalachian summits persisting as tundra much longer.
Tundra extended right to the Ice Sheet, forcing us to consider a transition of herd ranges rather than setting fixed boundaries. Beginning with the Deep South where calving occurred near Appalachian summits, the highest in this Chain, we think it likely that cows and calves descended spoke-like in early summer into this southernmost band of boreal forest, joining the bulls on the way. They may have migrated west from the Chain to northern Alabama and Mississippi, south to northern Georgia and southeast to South Carolina. The boreal band surrounding the calving ground was quite stable for centuries until the snowfields melted and boreal forest invasion forced safe calving north. A band of tundra reached southwest across eastern Tennessee where the Cumberland Plateau now exists. From 16K to 15K, its steep northeast-facing tundra slopes led early summer cows and calves down to their taiga winter range, with the reverse in spring when pregnant cows crossed this tundra strip to cross another taiga band to calve on tundra slopes.
By 14K, the snowfields had melted but the Appalachians remained tundra and pregnant cows continued to cross the Cumberland Plateau in early spring and early summer. Perhaps the southern arc of boreal forest continued to serve herds migrating semi-circularly and spoke-like around the southern extent of the Appalachians. But further north they migrated east and west on both sides of the Chain, the western herds crossing the still extant lobe extending southwest from the huge extent of tundra further north. It is possible that small herds to the north existed entirely on tundra, but this would depend upon the availability of ground lichen and shrubs, such that I believe they may not have lived right alongside the Ice Sheet or its meltwater lakes. From 15K to 12K, this tundra lobe paralleling the Chain was crossed by migrating herds on the west side, the east side having only taiga, due to Atlantic warming effects. The Appalachians were now taiga-covered and caribou date about 11.5K at Saltville in Virginia and Buckle’s Bog 115 km away.
At 11K, the Appalachian tundra continues to support calving grounds in a wide spoke-like arc like that in the south, emanating from the Appalachians from northeast to southwest completely across Pennsylvania. But it is now north-facing from Maine southwest across the rest of New England and northern New York and west to Ohio, Indiana and Illinois. This 3-layered arc of boreal forest, taiga and tundra is separated from the Great Lakes by a tundra band, its full extent from the Maritime Provinces across central Ontario and northern Michigan to Minnesota just as Lake Superior (Glacial Lake Minong) appeared. While Appalachian calving continued into Maine, calving grounds were also relocated in remote tundra locales near my South-Central Entrance (Niagara) to Ontario and in southern Michigan near my Southwest Entrance near Detroit-Windsor. We think it likely that calving grounds were also initiated at both ends of these bands in Wisconsin and Maine.
In concert with palaeoclimate and fluted points are artifacts of toolstone in sites that are traceable to the toolstone source. Using existing site and toolstone maps, plus many site reports, we prepared a table of about 100 fluted point sites where toolstone was known. Included were distance and direction from which a toolstone was taken to a site. We attempted to quantify toolstone types and locations, hoping to detect hunter movement north through a drop in number of southern toolstone pieces with a rise in pieces from northern sources. Maps with arrows from the toolstone source to each site were generated using a GIS from the table described above. Except for a few east-west arrows along the south shore of Lake Erie, all arrows followed a northeast-southwest line, the usual direction of herd movement. Some site clusters like those around Shoop in Pennsylvania had northeast-southwest arrows pointing in both directions, likely indicating relinquishment of southern cherts like Normanskill and initiation of Munsungan and Lake Champlain cherts as hunters moved north.
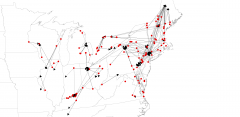
ENA toolstone sources to sites using arrows (Krist)
Relinquishment of southern toolstone sources and initiation of northern sources also occurred along my Southwest Entrance (Detroit-Windsor) where Michigan’s Bayport and Ohio’s Flint Ridge and Upper Mercer chert were imported to southwestern Ontario and relinquished as nearby Kettle Point and more northern Fossil Hill chert at Collingwood were adopted. This process also happened at my South-Central Entrance (Niagara Peninsula), where Pennsylvania Jasper and eastern New York Normanskill cherts were added to its local Onondaga and Haldimand cherts. All of the above discarding of southern toolstone sourcing and obtaining of new northern toolstone sources was integral to herd-following into southern Ontario.
1All dates are uncalibrated.